Oncology, Local Invasiveness - Oncologia, Invasività locale
Introduction
In this new article on the oncology series we are going to see what is the capacity of the tumor cell to become invasive, what mechanisms are modified and used to make colonization phenomena occur in tissues.
Local Invasiveness
The term local invasiveness refers to the ability of the tumour to invade the surrounding tissues, which can lead to metastasis.
One of the aspects that gives cancer cells this ability is the loss of contact inhibition, a mechanism that in physiological conditions gives healthy cells the ability to stop migrating and proliferate when they come into contact with each other.
In healthy cells, this mechanism is mediated by specific surface receptors, the caderins (in particular caderin-E, epithelial caderin), which allow two homotypical cells to recognize and bind together.

The extracellular portion of the caderine recognizes the identical domain expressed by the adjacent cell, so two caderines bind through the same identical molecular structure through a calcium dependent process, so the caderines are calcium dependent receptors**. When the recognition of the adjacent caderine takes place, at intracellular level the receptors recruit a multi-protein complex in which a class of proteins play a very important role. These are the catenins, which are organized at the level of the cytoplasmic portion of the receptor and act as a bridge between the receptors and the actin beams of the cellular cytoskeleton, thus bringing a certain rigidity to the cytoskeleton and to the cell itself, thus preventing their proliferation and migration.
Another important chain is the β-catenin, which also has another biological property that is to be a transcriptional factor, acting at the nuclear level causing cell proliferation. Therefore we can say that the engagement of caderins, determining the recruitment of chains, inhibits proliferation and movement, on the one hand because it engages the cytoskeleton, and on the other hand also because it sequester the β-catenin preventing it from going into the nucleus.
Many epithelial type tumors have a problem in homotypical recognition.
In many tumors, for example, caderin-E is down-regulated at the epigenetic level, so its gene is silenced, i.e. it is not expressed and therefore there is no contact inhibition.

Another factor related to caderine-E is its loss of expression and the acquisition of the expression of caderina-N (mesenchymal). This means that not only the tumor cell loses homotypic recognition, but it acquires other forms of recognition that allow it to interact with other cell types and in particular with mesenchyme cells. In addition, it should be remembered that, unlike the caderine E - caderine E interaction (very strong), the interaction of caderine N, like all interactions at the mesenchymal level, is much more lax so it may interact with mesenchymal cells but is not blocked or stopped. The ability to recognize mesenchymal cells and interact with them therefore facilitates the process of invasion and penetration of tumor cells within the stroma.
Together with caderine-E mutations there may also be mutations related to the β-catenins which may undergo a process of over-expression and therefore in addition to binding to the cytoplasmic tail of the receptors may go into the nucleus and lead to cell proliferation.
Epithelium-Mesenchymal Transition
The mutations observed so far are part of a large group of phenomena called epithelium-mesenchymal transition (EMT). EMT is a process whereby cells of epithelial nature acquire phenotypic traits specific to mesenchymal cells. In fact, they begin to express a whole series of proteins that are typical of mesenchymal cells, while they lose the expression of proteins that are typical of epithelial cells. Examples of some of these phenomena are:
-Loss of caderine-E typical of the epithelium and acquisition of caderina-N;
-Loss of cytoskeletal proteins typical of the epithelium and acquisition of vimentins typical of the mesenchyme;
-Loss of the typical polarity of epithelial cells to assume the characteristic apolarity of mesenchymal cells;
-Powering down a packet of epithelial cell gens;
-Shape: the epithelial cell is cuboid while the mesenchymal cell is more elongated;
-Increase in motility from epithelial cell to mesenchymal cell;
-Increased invasive capacity linked to the fact that mesenchymal cells are able to produce lithic enzymes capable of degrading the stroma;
-They begin to secrete intermediate filaments such as vimentin;
-They begin to secrete extracellular matrix proteins such as fibronectins (which the epithelium does not do);
-They begin to express ** integrinic receptors** other than those expressed by epithelial cells.

However, this EMT phenomenon does not occur at the level of all cancer cells, but mainly at the level of those cells that are in contact with the stroma.
All this modification of gene expression is coordinated through the expression of particular transcriptional factors that regulate the epithelium-mesenchymal transition by coordinating gene packages. Among the most important are SNAIL, SLUG, TWIST.
In turn, these transcriptional factors are activated as a result of mutations of certain genes in the tumor cell that cause this mechanism to be activated, or this process may be influenced by environmental factors.
In fact, it has been seen that a series of growth factors/cytokines produced by stromal cells (fibroblasts, macrophages, inflammation cells) intervene in this way and are able to ignite the phenomenon of epithelium-mesenchymal transition at local level; therefore they act on cells in the vicinity without genetically modifying them (epigenetic alteration).
The tumor cell, thanks to the help given by the stromal component, is able to elaborate an extremely effective proteolytic machinery that gives the tumor cell the ability, besides adhering and interacting with the extracellular matrix, also to "create a road". Among the most important lytic enzymes are the metalloproteases (collagenases), enzymes specific for different types of collagen that perform a single proteolytic cut in the collagen molecule so that the triple helix begins to unroll, showing the peptide bonds and becoming susceptible to the action of common proteases.
Another very important protease is the plasminogen, which, as with all proteases, is synthesized as a proenzyme and must meet a proteolytic cut to be activated.
Stromal cells produce two types of receptors for plasminogen activation:
the receptor for the urokinasic plasminogen activator (uPAR) (located on the migration front of tumor cells) which, by binding with the precursor pro-uPA (again produced by the stroma), activates it by transforming it into uPA. This protease activates the plasminogen, which is a broad spectrum protein produced by the liver, by proteolytically cutting the protein that becomes plasmin. The fundamental role of plasmin is to activate the metalloproteases so that they can perform the degradation action at the collagen level. In addition, plasmin is able to activate TGF-β, which promotes the production of metalloproteases by tumor cells.
the receptor for the activation of the tissue type plasminogen (tPA), activates the plasminogen in the same way as uPAR but in this case it serves to degrade fibrin clots.
There is also another type of metalloprotease which are the transmembranal metalloproteases which are proteases that have the function of degrading the matrix, but have the peculiarity of being permanently anchored to the cell surface.
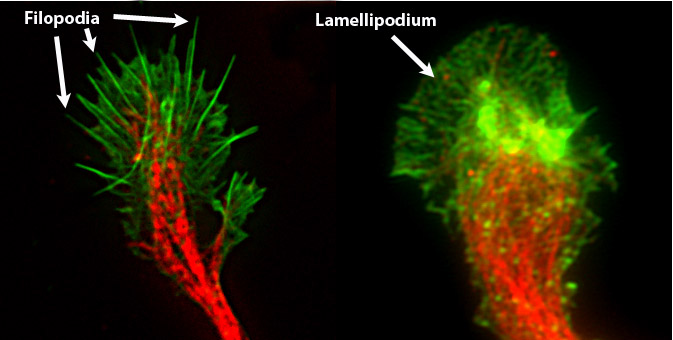
The tumor cell has to move and does so through various mechanisms, the most important of which concerns the emission of a pseudopodo (lamellipod) which interacts with the external environment and acts as a traction point to drag the cell (amoeboid movement). This movement involves a reorganisation of the cytoskeleton on the migration front and this interacts with the external environment thanks to specific receptors, which are the integrins, which act as a bridge between the surrounding environment and the intracellular environment. The lamellipodio must therefore be hooked to the substrate to interact with the matrix and does so using the appropriate integrins. Once attached, the cell is able to move and this involves a series of reorganizations of the cytoskeleton. The lamellipod is therefore characterized by the presence of "spines" which are the philipods which are cytoskeletal protrusions that touch the environment and transmit the signal of the conditions outside the intracellular environment, so as to facilitate the movement

Introduzione
In questo nuovo articolo sulla serie di oncologia andremo a vedere quella che è la capacità della cellula tumorale di diventare invasiva, quali meccanismi vengono modificati e utilizzati per far si che avvengano fenomeni di colonizzazione nei tessuti.
Invasività Locale
Con il termine invasività locale ci si riferisce alla capacità del tumore di invadere i tessuti circostanti favorendo un’eventuale successiva invasività distale che può portare a metastasi.
Uno degli aspetti che conferisce alle cellule tumorali tale capacità è data dalla perdita dell’inibizione da contatto, meccanismo che in condizioni fisiologiche conferisce alle cellule sane la capacità di smettere di migrare e proliferare quando vengono a contatto l’una con l’altra.
Nelle cellule sane questo meccanismo è mediato da recettori di superficie specifici, le caderine (in particolare la caderina-E, caderina epiteliale), che permettono a due cellule omotipiche di riconoscersi e legarsi tra loro.
La porzione extracellulare della caderina riconosce lo stesso identico dominio espresso dalla cellula adiacente, quindi due caderine si legano attraverso la stessa identica struttura molecolare attraverso un processo dipendente dal calcio, quindi le caderine sono dei recettori calcio dipendenti. Nel momento in cui avviene il riconoscimento della caderina adiacente, a livello intracellulare i recettori reclutano un complesso multi-proteico in cui giocano un ruolo molto importante una classe di proteine che sono le catenine, le quali si organizzano a livello della porzione citoplasmatica del recettore e fanno da ponte tra i recettori e i fasci di actina del citoscheletro cellulare, portando così una certa rigidità al citoscheletro e ad alla cellula stessa, impedendone così la proliferazione e migrazione.
Altra catenina importante è la β-catenina, la quale ha anche un’altra proprietà biologica che è quella di essere un fattore trascrizionale, agendo a livello nucleare determinando proliferazione cellulare. Quindi si può dire che l’ingaggio delle caderine, determinando il reclutamento delle catenine, inibisce la proliferazione e il movimento, da un lato perché ingaggia il citoscheletro, e dall’altro lato anche perché sequestra la β-catenina impedendole di andare nel nucleo.
Tantissimi tumori di tipo epiteliale hanno un problema a livello di riconoscimento omotipico.
In molti tumori per esempio la caderina-E è down-regolata a livello epigenetico, quindi il suo gene viene silenziato, ovvero non viene espresso e quindi non c’è inibizione da contatto.
Un altro fattore legato alla caderina-E è la sua perdita di espressione e l’acquisizione dell’espressione della caderina-N (mesenchimale). Questo significa che non solo la cellula tumorale perde il riconoscimento omotipico, ma acquisisce altre forme di riconoscimento che permettono di interagire con altri tipi cellulari e in particolare con cellule del mesenchima. Inoltre bisogna ricordare che, a contrario dell’interazione caderina E - caderina E (molto forte), l’interazione della caderina N, come tutte le interazioni a livello mesenchimale, è molto più lassa quindi può sì interagire con le cellule mesenchimali ma non per questo viene bloccata o arrestata. La capacità di riconoscere le cellule mesenchimali e interagire con loro facilita quindi il processo di invasione e penetrazione delle cellule tumorali all’interno dello stroma.
Insieme alle mutazioni delle caderine-E ci possono anche essere mutazioni relative alle β-catenine che possono subire un processo di over-espressione e quindi oltre a legarsi alla coda citoplasmatica dei recettori possono andare nel nucleo e determinare proliferazione cellulare.
Transizione Epitelio-Mesenchimale
Le mutazioni fino ad ora osservate rientrano in un grande gruppo di fenomeni che stanno sotto il nome di transizione epitelio-mesenchimale (EMT). La EMT è un processo tale per cui cellule di natura epiteliale acquisiscono tratti fenotipici propri delle cellule mesenchimali. Esse infatti iniziano a esprimere tutta una serie di proteine proprie delle cellule mesenchimali, mentre perdono l’espressione di proteine proprie delle cellule epiteliali. Esempi di alcuni di questi fenomeni sono:
-Perdita di caderina-E tipica dell’epitelio e acquisizione della caderina-N;
-Perdita di proteine del citoscheletro proprie dell’epitelio e acquisizione di vimentine proprie del mesenchima;
-Perdita della polarità tipica delle cellule epiteliali per assumere la caratteristica apolarità delle cellule mesenchimali;
-Spegnimento di un pacchetto di geni relativo a cellule epiteliali;
-Forma: la cellula epiteliale è cuboide mentre quella mesenchimale è più allungata;
-Aumento della motilità passando dalla cellula epiteliale alla cellula mesenchimale;
-Aumenta la capacità invasiva legata al fatto che le cellule mesenchimali sono in grado di produrre enzimi litici in grado di degradare lo stroma;
-Iniziano a secernere filamenti intermedi come la vimentina;
-Iniziano a secernere proteine proprie della matrice extracellulare come fibronectine (cosa che l’epitelio non fa);
-Cominciano a esprimere recettori integrinici diversi da quelli espressi dalle cellule epiteliali.
Questo fenomeno di EMT non avviene però a livello di tutte le cellule tumorali, ma avviene soprattutto a livello di quelle cellule che sono a contatto con lo stroma.
Tutto questa modificazione di espressione genica viene coordinata attraverso l’espressione di particolari fattori trascrizionali che regolano la transizione epitelio-mesenchimale coordinando pacchetti di geni. Tra i più importanti abbiamo SNAIL, SLUG, TWIST.
A loro volta questi fattori trascrizionali vengono attivati a seguito di mutazioni di alcuni geni nella cellula tumorale che fanno sì che questo meccanismo venga attivato, oppure tale processo può essere influenzato da fattori ambientali.
Si è infatti visto che una serie di fattori di crescita/ citochine prodotte da cellule stromali (fibroblasti, macrofagi, cellule dell’infiammazione) intervengono in questa via e sono in grado di accendere il fenomeno della transizione epitelio-mesenchimale a livello locale; quindi agiscono sulle cellule poste nelle vicinanze senza modificarle geneticamente (alterazione epigenetica).
La cellula tumorale grazie all’aiuto dato dalla componente stromale è in grado di elaborare un macchinario proteolitico estremamente efficace che conferisce alla cellula tumorale la capacità, oltre ad aderire ed interagire con la matrice extracellulare, anche di “crearsi una strada”. Tra gli enzimi litici più importanti ricordiamo le metalloproteasi (collagenasi), enzimi specifici per i diversi tipi di collagene che eseguono un unico taglio proteolitico nella molecola di collagene in modo tale che la tripla elica cominci a srotolarsi, mostrando i legami peptidici e diventando suscettibile all’azione delle proteasi comuni.
Un’altra proteasi molto importante è il plasminogeno, il quale, come accade per tutte le proteasi, viene sintetizzato come proenzima e per essere attivato deve andare incontro a un taglio proteolitico.
Le cellule stromali producono due tipologie di recettori per l’attivazione del plasminogeno:
il recettore per l’attivatore del plasminogeno di tipo urochinasico (uPAR) (localizzato sul fronte di migrazione delle cellule tumorali) che legandosi con il precursore pro-uPA (prodotto ancora una volta dallo stroma) lo attivano trasformandolo in uPA. Questa proteasi attiva il plasminogeno, che è una proteina ad ampio spettro prodotta dal fegato, tagliando proteoliticamente la proteina che diventa plasmina. Il ruolo fondamentale della plasmina è quello di andare ad attivare le metalloproteasi in modo che queste possano eseguire l’azione di degradazione a livello del collagene. Inoltre la plasmina è in grado di attivare il TGF-β, la quale favorisce la produzione di metalloproteasi da parte delle cellule tumorali.
il recettore per l’attivazione del plasminogeno di tipo tessutale (tPA), attiva il plasminogeno allo stesso modo dell’uPAR ma in questo caso serve per degradare i coaguli di fibrina.
Esiste anche un’altra tipologia di metalloproteasi che sono le metalloproteasi transmembranali che sono proteasi che hanno la funzione di degradare la matrice, ma hanno la peculiarità di essere ancorate stabilmente alla superficie cellulare.
La cellula tumorale si deve muovere e lo fa attraverso vari meccanismi, il più importante dei quali riguarda l’emissione di uno pseudopodo (lamellipodio) che interagisce con l’ambiente esterno e fa da punto di trazione per trascinare la cellula (movimento ameboide). Questo movimento prevede una riorganizzazione del citoscheletro sul fronte di migrazione e questo interagisce con l’ambiente esterno grazie a specifici recettori, che sono le integrine, le quali fanno da ponte tra l’ambiente circostante e l’ambiente intracellulare. Il lamellipodio si deve quindi agganciare al substrato per interagire con la matrice e lo fa mediante le opportune integrine. Una volta che si è attaccata, la cellula è in grado di muoversi e ciò comporta una serie di riorganizzazioni del citoscheletro. Il lamellipodio è quindi caratterizzato dalla presenza di “spine” che sono i filipodi che sono delle protrusioni citoscheletriche che toccano l’ambiente e trasmettono il segnale delle condizioni esterne all’ambiente intracellulare, in modo da favorire il movimento.
Fonti/Sources
Immagini/Pictures
{kind=link}
{kind=link}
{kind=link}
{kind=link}