Organogenesis
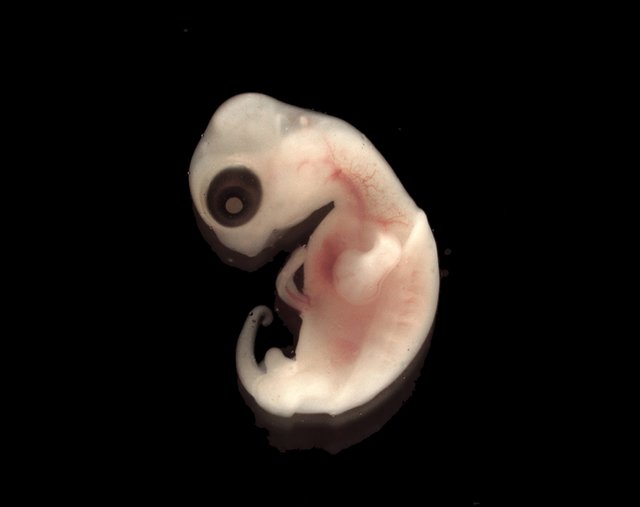
Though animals are dramatically diverse, the basic mechanisms of their development are similar across groups.
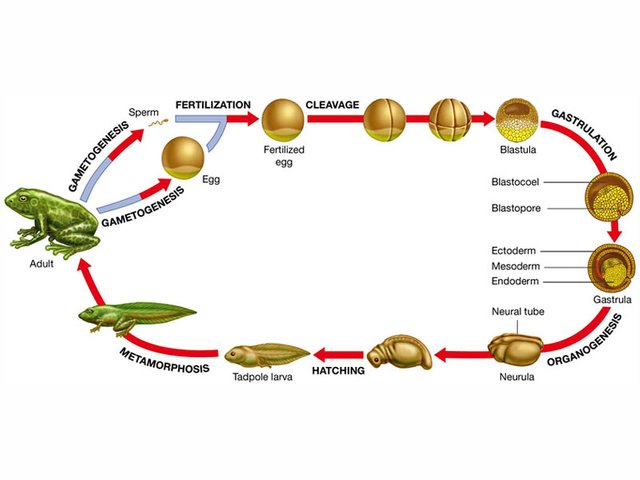
Today we examine the four earliest stages of development fertilization, cleavage, gastrulation, and organogenesis and outlines the processes involved in each. After fertilization, each stage involves a set of simple processes repeated over and over to produce the great diversity of cell types and structures found in a complex multicellular organism, and ultimately the diversity we observe across different kinds of organisms.
The specific processes and patterns of development differ greatly among different groups of animals, but all animals share much in common in the early stages of development.
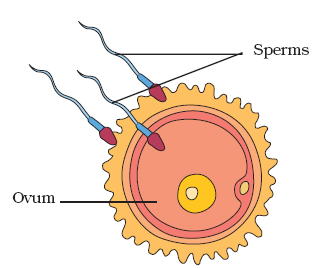
The first step in the development of a sexually reproducing organism is fertilization—the creation of a zygote from the fusion of a female gamete and a male gamete. Fertilization is more complex than the random joining of gametes; the ability of a sperm to penetrate an egg depends on an intricate set of cell-signaling mechanisms. Fertilization initiates a series of events that cause the zygote to become inherently asymmetric—that is, to have an asymmetric distribution of protein and messenger RNA (mRNA) in the cytoplasm. In some organisms, this asymmetry is visually apparent because it is associated with an equally asymmetric distribution of yolk that can be observed under a microscope or, sometimes, with the naked eye. To classical embryologists, the yolk-containing end of a zygote is known as the vegetal pole, while the end with less yolk is known as the animal pole. This distinction leads to a useful picture of a zygote as a globe with north and south poles. Though the zygotes of many animals do not contain visible yolk, they do have an asymmetric distribution of protein and mRNA. This asymmetry is one of the sources of information that may cause the zygote’s daughter cells to diversify.
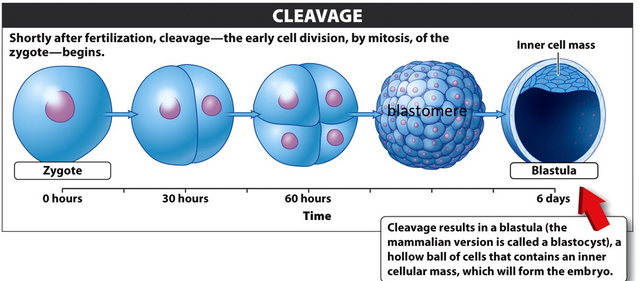
After fertilization, the zygote undergoes a rapid series of cell divisions called cleavage.
Cleavage is simply a form of mitosis; the resulting daughter cells are called blastomeres. The cell divisions that occur during cleavage follow a very orderly pattern with respect to cell orientation. As in normal mitosis, a dividing cell separates along the plane formed by the chromosomes during metaphase (the metaphase plate). If a zygote is pictured as a globe and the first division of cleavage divides it along a longitudinal axis, the two blastomeres will have the same north-south orientation as the zygote. However, blastomeres also may divide along the “equator” of their globes (along an equatorial axis), producing daughter cells with a different orientation. The details of which cells divide along which axis are unimportant, and different groups of animals follow different patterns. The important point is that the first series of cell divisions during cleavage does follow a specific pattern of orientations.
This specific pattern of cell orientations is important during cleavage because, depending on their orientations, blastomeres may or may not preserve the asymmetry of the parent cell. The regular pattern of orientations in the first several divisions of cleavage establishes another level of asymmetry in the developing organism, with some blastomeres having the same relative arrangement of proteins and mRNAs (cytoplasmic determinants) as their parents and other blastomeres becoming distinctly different. Another characteristic of cleavage is that there is no net increase in the volume of cytoplasm between cell divisions. The cells, therefore, become progressively smaller with each division, and there is little or no increase in the size of the embryo. Because there is no growth, cell division also occurs at a very rapid rate.
One final feature of cleavage is that, in general, the genes of the developing organism are not expressed. DNA is replicated but not transcribed, which suggests that cleavage must be controlled by the proteins and mRNAs—the cytoplasmic determinants—found in the zygote. After some number of cleavages has occurred, the developing organism turns into a solid ball of small cells called a morula. A fluid-filled cavity then develops inside the ball of cells, forming a hollow ball of cells called a blastula. The inner space of this hollow ball is called the blastocoel, and the sheet of cells that surrounds it is called the blastoderm.
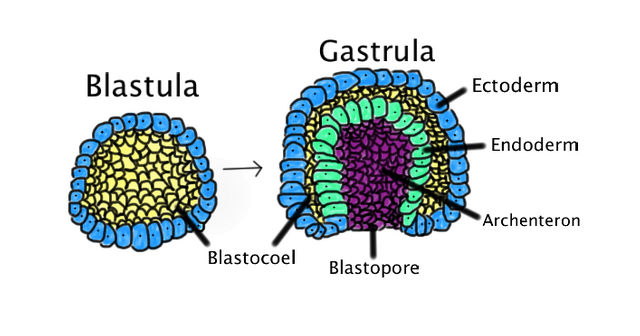
The next key stage in early development is called gastrulation.
During which the cells of the developing organism fold into more complex shapes and begin to assume different developmental fates. Gastrulation varies even more among groups of animals than fertilization or cleavage, but gastrulation in all animals shares a common set of mechanisms. During gastrulation, morphogenesis becomes particularly important.
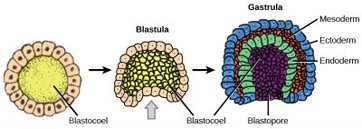
Gastrulation involves three major morphogenetic phases.
- First, some cells move from the surface of the blastula to its interior.
- Second, as a consequence of this migration, three distinct layers of different kinds of cells are formed in the embryo.
- Third, by the end of gastrulation, the blastula has become an embryo, with an internal digestive cavity that is connected to the outside and that defines the anterior-posterior axis of the animal.
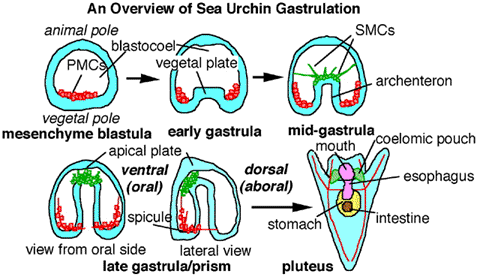
The sea urchin provides a convenient model for describing gastrulation.
Gastrulation in sea urchins begins at the vegetal pole, where the cells flatten and form the vegetal plate. A number of cells from the vegetal plate (called mesenchyme cells) detach from the blastoderm and enter the blastocoel as loose cells. The cells in the vegetal plate change shape and begin to buckle inward. The inward-extending sheet of vegetal plate cells forms a deep pouch inside the blastocoel called the archenteron, which is connected to the outside by a hole (the blastopore). The closed end of the archenteron eventually extends all the way to the outer wall of cells on the other side and forms a second opening, creating a tube that will become the sea urchin’s gut.
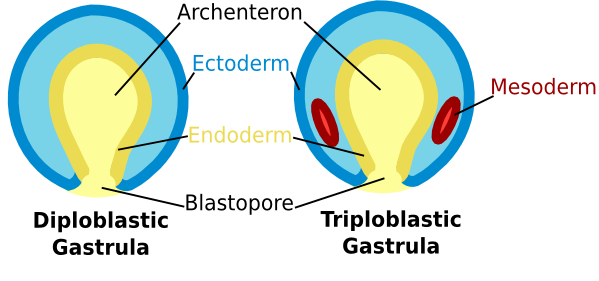
The final product of gastrulation is an embryo with a primitive gut and three distinctive cell layers.
The ectoderm is the outermost layer; it is derived from those cells of the original blastoderm that remained on its outside through the entire process. The innermost layer is the endoderm; it is the layer formed when cells at the vegetal plate extended to the inside of the blastoderm as the archenteron formed. Finally, there is a third layer of cells, called the mesoderm, located between the ectoderm and endoderm. The cells of the mesoderm are derived from mesenchyme cells that detached from the original hollow ball and migrated into the blastocoel.
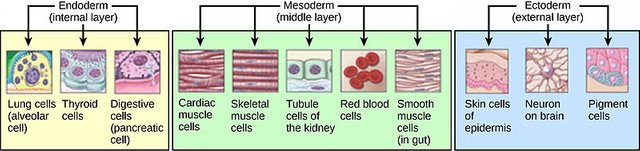
These three layers (also called the embryonic germ layers) give rise to distinctly different kinds of tissues in the adult animal. In vertebrates, the ectoderm obviously gives rise to the outer layer of the skin (or epidermis) and derivatives of the epidermis, such as hair and nails. It also gives rise to much of the nervous system, the lining on the inside of the mouth and the cornea and lens of the eyes, and many glands found in the skin. The mesoderm gives rise to the skeletal system, muscles, circulatory system, gonads, and some internal organs, such as the kidneys and the heart. The endoderm gives rise to the inner linings of the digestive and respiratory systems, as well as other internal organs, such as the liver and pancreas.
>
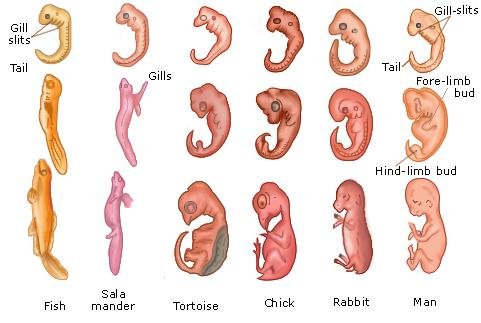
The formation of rudimentary limbs, appendages, and organs that follows gastrulation is referred to, in general, as organogenesis.
The specific events that occur during organogenesis differ radically from animal group to animal group, giving rise to the enormous diversity in body structure among different kinds of animals. In spite of this diversity, all these morphogenetic movements, to a large extent, simply represent the folding and bending of the three embryonic tissue layers established during gastrulation. The establishment of internal structures, such as organs, and external structures, such as limbs, involves a series of invaginations (“inpocketings”) and evaginations (“out-pocketings”) of the different germ layers.
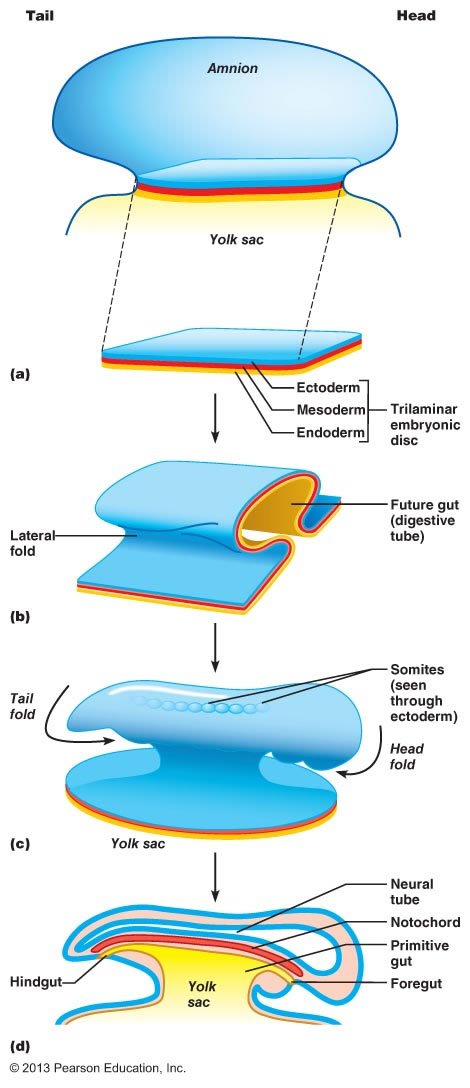
To picture the developments of organogenesis, imagine slicing an embryo from “north” to “south”—this would be equivalent to slicing crosswise through a hollow tube that has three concentric layers running along its length. In organogenesis, different layers at different points along the length of this tube are bended and folded inward to form invaginations or outward to form evaginations. For example, in vertebrate animals, one of the earliest structures to form is an additional tube running along most of the length of the embryo that will eventually become the spinal cord and brain. This tube is called the neural tube, and it is formed as an invagination of the ectoderm.
The important point is that the essentially simple process of morphogenetic movement is repeated over and over again, with different variations and in different places, to create the rudiments of all the organs and appendages of a complex animal body.
Sources:
Image Credits:
https://www.nsf.gov/news/mmg/media/images/reptile_hearts4_h1.jpg
http://biology.kenyon.edu/courses/biol114/Chap12/FG18_05.JPG
http://www.ekshiksha.org.in/images_08.09ReproductionInAnimals/ferilization.png
https://classconnection.s3.amazonaws.com/929/flashcards/1460929/png/cleavage1336950416807.png
https://upload.wikimedia.org/wikipedia/commons/d/d9/Blastula.png
http://study.com/cimages/multimages/16/0blastulax.jpg
http://worms.zoology.wisc.edu/urchins/SU_gast_over.gif
http://study.com/cimages/multimages/16/gastrula1.png
https://upload.wikimedia.org/wikipedia/commons/thumb/5/52/Germ_layers.jpg/800px-Germ_layers.jpg
https://mrkubuske.files.wordpress.com/2014/03/vertebrate-embryos.jpg
http://classes.midlandstech.edu/carterp/Courses/bio211/chap28/figure_28_10_labeled.jpg
Bird [Giphy](https://giphy.com/gifs/blue-5lDik5jPpRw2Y)
BTC: 1JQWNxziArfaCtxsP3tUQ4VSTBY9jresmt 

nice post. thank you for explaining clevage. whole new light. haha
I had considered using different images in that section. ;)
Great post. I love how you thoughtfully bring out our connectedness to all living things. 5 kudos !
Nice job PJ. Keep it up!