The "cell computer", an organelle of microtubules, is also replicating during cell division
"Boveri (1901) described the centrosome as "an autonomous permanent organ of the cell - the dynamic center of the cell - the true division center [whose] division creates the center of the daughter cell" (from Mazia 1984). " [1]
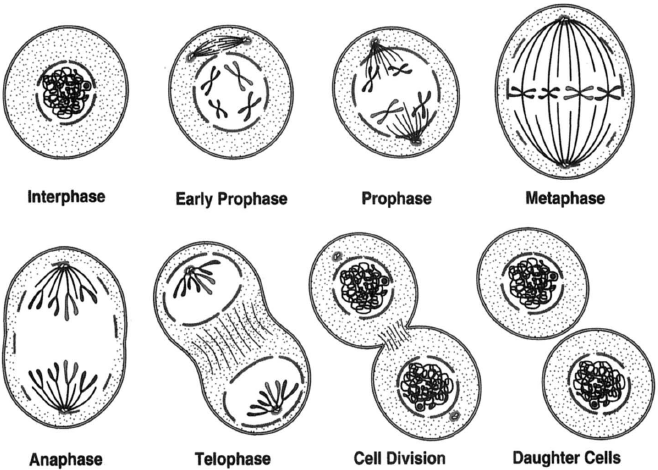
Cell division is organized around that the cell forms a bipolar spindle during prophase, and to achieve this, the centrosome must be replicated accurately during interphase.[2] There are parallels with DNA replication, centrosome duplication and DNA replication are initiated at the same time in the cell cycle, and, like DNA replication, centrosome duplication can be considered to be semiconservative, with each centrosome receiving one old centriole and one new centriole. [3]
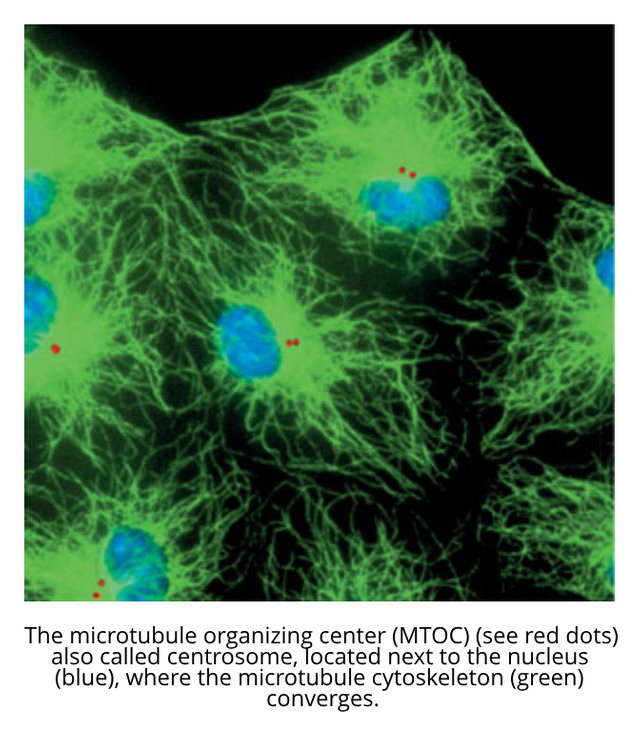
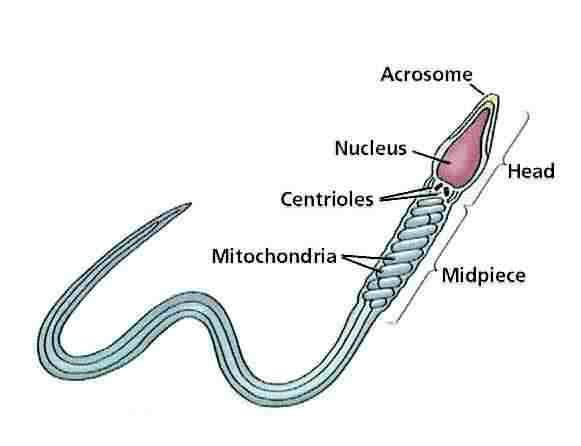
The original theory of the centrosome as the 'reproductive organ' of the cell provides a logical explanation of the mitotic poles and the accuracy of cell division. Ontological evidence for the reality of centrosomes came out of the discovery of the nature of fertilization. The spermatozoon 'injects' into the ovum a nucleus and a centrosome. The injected centrosome is the ancestor of all the centrosomes in all of the cells of the developing organism.[4, 5]
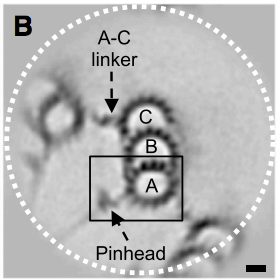
What is the structure of a centriole, prediction from information theory
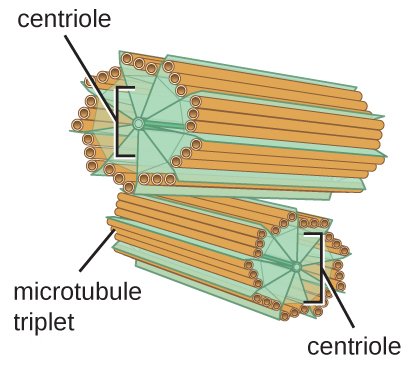
With a 13 + 10 + 9 proto-filament structure, reflecting two 13 filament MTs on opposite sides of a 14 filament MT, a microtubule triplet would have a circumference of exactly 8 memory blocks. In this electron microscopy image from 2013 and Guichard et al[6], the A-tubule appears smaller than the B-tubule because it is, and the C-tubule appears larger than the A-tubule because it conforms to the larger B-tubule (whereas A does not adapt to B, B adapts to A. )
That the C-tubule is one protofilament less than the B-tubule would also make the triplet microtubule symmetric, the A-tubule having 9 filaments towards the surface, the C-tubule as well, with the B-tubule in between, exposing 3 filaments to the outside.
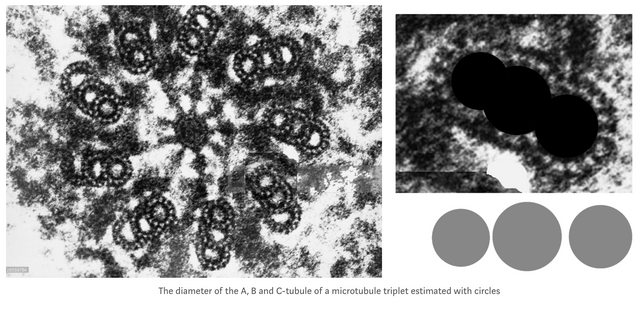
Since the 8 filaments between the A, B and C-tubule are "wasted" from an information theory perspective, there is also no need for a helical structure to fit the blocks onto the triplet microtubules (on isolated microtubules with 13 filaments, the 3 x 2 tubulin blocks need to form a coil to be able to fit. ) It was also shown in 1995 that the inner filaments are not tubulin, they are composed of tektin with a 48 nm axial repeat, 3x that of tubulin dimers.[7]

An estimate of the memory storage capacity of a centriole
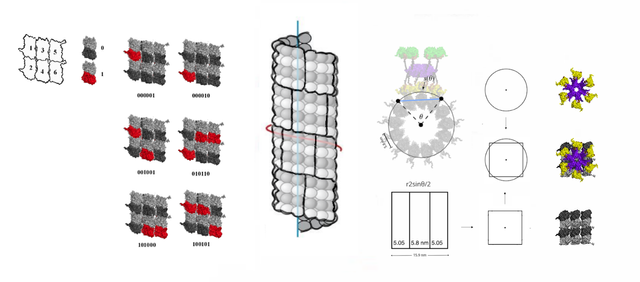
The centriole, a cylinder about 500 nm long composed of short lengths of microtubules, could store 31 memory blocks in height, 16 nm * 31 ≈ 500 nm (32 blocks if 512 nm high), and can fit 8 blocks around the circumference of each microtubule triplet, with a total of 8 * 9 * 31= 2232 blocks of data, 2232 * 6 * log2(3) = 2.65 kb, and 3^6 = 729 possible states per block, and 313392 = 4*106389 possible states in a single centriole, 326784 possible states per centrosome (two centrioles).
The centrosome as a reference homogenic field, to co-ordinate the cell computer and overall cell function
"Of course, the geometric similarities between the hypothetical detector and centrioles may be coincidental; and the postulated signals may never be found. On the other hand, they have not been looked for, either. Should such signals exist - be they electric currents, infrared light emissions corresponding to molecular vibrations, or others - entirely new aspects of the control of intracellular processes would emerge. " [8]
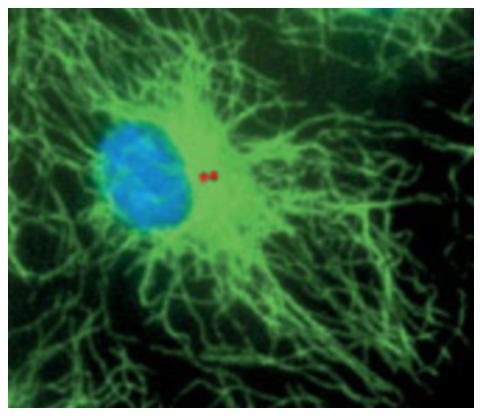
The centrosome (red dots in image below) as a BIOS of the cell, with a state of 5.31 kb, could be the center of the electromagnetic field of the cell[9], and amplify it, and emit a signal that forms a reference field for the cell computer, a medium where information can be weaved into coherent and meaningful ideas.
What is the total storage capacity of microtubules?
Danko Georgiev estimates that there are 1.3×10⁹ tubulin dimers in adult pyramidal neurons[10], and as a trinary system with three possible states per tubulin (0, 1 and 2), log2(3) * 1.3*10⁹ = 256.75 mb. With 10–100 trillion cells, even a tiny data storage capacity of 10 mb per cell, 5×10⁷ tubulin dimers, is 10 mb * 10 trillion cells = 10 * 10¹³ = 10¹⁴ mb = 100 exabytes.
An exponential rate for spiraeum replication via tree-like microtubule nucleation
Branching nucleation is an ideal mechanism for replicating the spiraeum during the prophase of mitosis, new daughter microtubules can form at the side of existing parent microtubules, resulting in the accelerated formation of the total length of copied microtubules within the parallel microtubule structure of the mitotic spindle.
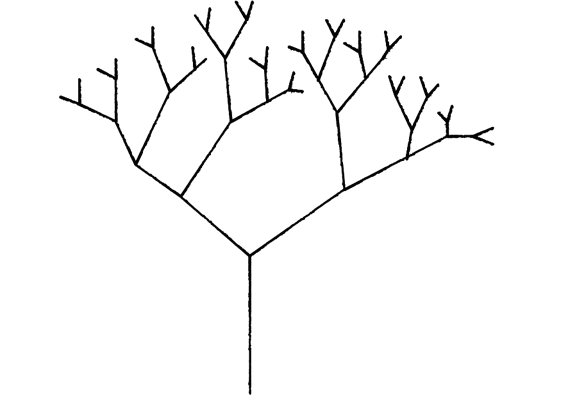
With 30 nm/s extension rate of microtubules, each branch forms two coil turns per second, 9 blocks of memory which can be seen as 9 append-only inputs per second. If prophase is 2 hours, 7200 seconds * 9 blocks per second = 64800 memory blocks copied during prophase, 77 kilobytes per branch within the mitotic spindle, a spindle apparatus with 100 branches could at a very low growth rate of 30 nm/s replicate 7.7 mb in just 2 hours.
Synapses
Centrosome replication in somatic cells: the significance of G1 phase (2000)
Centrosome Duplication (2001)
Paternally contributed centrioles exhibit exceptional persistence in C. elegans embryos (2015)
At least one of the protofilaments in flagellar microtubules is not composed of tubulin (1995)
Remarks on the number of tubulin dimers per neuron and implications for Hameroff-Penrose
Orch OR (2009)Does the geometric design of centrioles imply their function? (1981)
Cell Division: How centrioles acquire the ability to reproduce (2017)
Great and excellent indepth details on that beyond imagination